Background
In the last
section, we’ve seen how MGAs may be used to develop a model. However, when
using them, it is quite possible you may get a plot of fitness of creatures in
the population as follows (apologies for its ugly printscreen nature; it comes
from a quick snap into a digital logbook, as is the case with most figures in this section):

On the
x-axis is the fitness component corresponding to action potential morphology, on
the y-axis is the compound value aggregating other criteria. You can see the
tradeoff between the two dimensions easily. In case you’re wondering about what
is the green rectangular object, it shows the zone that corresponds to what I
thought would be a reasonably good solution at the time. Of course, nothing
inside, nor even close :( This means one of two things; either there is
a model parameter that we are missing in the creature DNA (good luck finding it
among the hundreds of possible numbers in the model), or there is a structural
problem in the model – an important current may be missing, or the formulation
of a current is problematic. Below, we’ll
go over a concrete example that I encountered in model development of what one
may do in the case where an automated optimizer fails to find a suitable
solution. The abbreviation I selected for this process is CSOMSKY (Case Study Of Manual Searching, King in Yellow [1]).
I note that
the plots used in this section are, unless specified otherwise, based on an
intermediate development version of the model and are not representative of the
final model version.
Diagnostics
The first
step is to get proper diagnostics. The fitness function in Matlab returns only
the fitness number[s], in our case two fitness dimensions. However, as
mentioned previously, one of the dimensions was in fact aggregating more
calibration criteria (calcium transient properties and response to INa+INaL,
ICaL, and IK1 partial blocks). It’s thus useful to make a
version of fitness function which also returns the calibration criteria in
isolation, before they are weighted-summed into a fitness component. It may be
also of use to return the traces of action potential, calcium transient, etc.
in case one wants to inspect them too. This diagnostic-fitness function may be
run after the MGA finishes, running it on creatures on the pareto front,
gaining more information about them.
First steps
What I
expected the most was to find a single calibration criterion that prevented
achieving an overall satisfactory fitness. If that was the case, I’d have
probably tried to find the difference between models which perform best and
worst at the calibration criterion, taking it further from there.
Interestingly,
the situation was different, and it was actually two criteria together causing
the problem: one requiring the calcium transient amplitude to reduce in response to 50% sodium blockade (blocking both INa and INaL), and the other requiring APD shortening in response to 50% ICaL block. Class A of creatures produced models which correctly reacted to sodium blockade, but incorrectly prolonged the action potential duration in
response to the block of ICaL. Class B of creatures was the
opposite, manifesting an incorrect reaction to sodium block, but a correct one
to the ICaL block. There was a clear tradeoff between the two
calibration criteria – some models were clearly good at one and horrible in the
other, and then also models that were quite balanced, being slightly bad in
both. For further investigation, I decided to focus on the
models clearly in class A or in class B.
As the next
step, I sought to find out what is the difference between the two classes;
given that most of our creatures’ DNA was current multipliers, this can be
worded as finding which currents differ mainly between the two populations [2].
The current most strongly and consistently different between creatures
belonging to class A and B was INaL, the late sodium current (with
class A having a high INaL).
Verification of the first
step
It might
still be very well possible that INaL is the main difference, but
not the causative factor. To verify the causal role, I took creatures of class
A (good sodium block, poor ICaL block) and reduced their INaL.
The creatures changed into class B, suddenly manifesting a poor reaction to
sodium block, but a good one to ICaL block. Thus, INaL
appears to be a causative factor. For completeness of information, I measured
the effect of INaL reduction in two ways which give slightly
different information:
- Pre-pacing a control cell for 1000 beats, followed by acute INaL reduction for 5 beats and measurement of the cell properties during these. This gives information about the direct electrophysiological effect of the current reduction. In this particular case, the acute change induced a clear change in behavior, suggesting a direct electrical influence of INaL on the phenomena of interest.
- Chronic INaL reduction, i.e., reduction of INaL already from the start of simulation, followed by pacing for 1000 beats. This was then compared to a cell paced for 1000 beats without the INaL reduction. If there was no major change in cell behavior with acute change (which we did see, as described above), but there was an effect with a chronic change, it would have suggested the role of INaL is about a long-term effect such as a change in ionic concentrations.
Another
approach that I did not use here, but that can be used to dissect causality, is
clamping of variables. Do you suspect that CaMKII phosphorylation of a channel
promotes EAD formation? Try to repeat the simulation with the said
phosphorylation (on that site only) clamped not to exceed a given threshold –
if that stops the EAD facilitation, it’s one more reason to think your
suspicion holds water.
Finding the culprit
We have
discovered that INaL plays a large role in the fork between good
behavior of sodium and ICaL blockade, however, this does not tell us
anything about the mechanism. At the same time, finding the mechanism is
critical, as that’s what can eventually lead us to an improvement.
I took a
model of class A as a starting point, trying to understand why its reaction to
ICaL block is poor, showing an APD prolongation. Starting with class
A model rather than class B was simply because a good reaction to sodium
blockade was one of the most important criteria to us – it is in fact where the
whole project on development of ToR-ORd started.
The AP
morphology of the selected intermediate model in control condition (blue) and
under ICaL block (red) looked as follows:

(x axis is time - last spike of many, hence the weird range, and y axis is the membrane potential)
For this part of investigation, I used a 100 % ICaL block to make the effect as clear as
possible. Subsequently, I plotted the currents in control and ICaL
block conditions, looking for main differences in these.
Unsurprisingly,
the largest difference in currents was in the reduction of ICaL with its full
block, but this would strongly shorten the APD if other factors were kept
constant, so it was certainly not the explanation. Rather, it was clear that
there are other factors which override this trend. The two currents changed
most prominently by the full ICaL block were increased INaL,
and reduced IKr, both contributing to the APD prolongation.
Culprit 1 – INaL
The INaL in
the two conditions (identical color coding to the previous plot) looked as follows:

The
increase in INaL amplitude following ICaL block could be due to changes in activation, inactivation, or driving force.
An inspection of state variables during an action potential revealed that the
dominant factor in the INaL increase is the increase in driving
force, which follows from lowered plateau potential under ICaL
block.
Is such an
increase in INaL plausible? In general, yes, the increase in driving
force is hard to argue against. However, one may wonder if there aren’t other
factors coming into play, linked to special gating properties of the Nav 1.5
channel mediating INa and INaL. Studies based on
single-pulse voltage clamp predicted that INaL would be shaped like
an extended triangle-like decay of INa; however, a recent-ish study (Horvath et al. 2013) employing an AP clamp technique suggests
it is not really the case (Figure 1A in this article), explaining the
difference via non-equilibrium gating (there are some very interesting papers
on this, e.g. (Clancy et al. 2003))[3].
If it is the case that INaL is generally small shortly after AP
upstroke as shown in the Horvath et al. paper, perhaps the effect of increased driving force is mitigated. However,
in the absence of data to decide what is happening with INaL under ICaL
block, it was hard to say what to do and I decided to switch focus to the considerably
reduced IKr.
Nevertheless,
at this stage, it dawned on me why INaL was at the core of the fork
between classes A and B in the population returned by the MGA. As described in
the ToR-ORd paper, a reduction in INaL reduces the calcium transient
amplitude; mainly via APD shortening and resulting smaller calcium influx via If ICaL. If INaL is generally high, its
reduction causes a more prominent APD shortening and thus a greater reduction
in calcium transient amplitude. This is why models with higher INaL
could manifest a net calcium transient amplitude reduction in response to 50/50
INa+INaL block. However, at the same time, given that INaL
is increased under ICaL block due to the increased driving force,
models with high INaL show a greater total increase in INaL
following an ICaL block, and thus manifest APD prolongation,
performing poorly on the criterion of ICaL block and APD shortening.
Despite the utility of understanding the fork between classes A and B, it was
pretty clear that yet again, understanding a problem does not mean it’s obvious
how to solve it.
Culprit 2 – IKr
The second
factor involved in the APD prolongation in response to ICaL block
was IKr; at the time, the current formulation was the same as in the O’Hara-Rudy (ORd) model. Following ICaL blockade, IKr was reduced considerably:

Again, this
reduction could be, in theory, due to changes in inactivation, activation, or
driving force. Inspection of state variables has shown it is the activation –
and it could be either a change in the steady-state value of activation, or the
time constant of reaching it. Plotting the steady-state values for the given
membrane potentials versus the actually achieved membrane potential showed a
marked difference, pointing to the time constant as the causative factor.
The reason
for the lack of IKr is described in our paper, but to briefly recapitulate it here, let’s look at the
plot of the fast and slow activation time constants of ORd model’s IKr
(to the left, taken from the original ORd article), and the action potential morphology of the final
ToR-ORd [4] as well as ORd (to the right, taken from our paper):

The
membrane potential during the early plateau in our model (both the intermediate
and final ToR-ORd) is around 20-23 mV, and this clearly corresponds to low
values of fast and slow IKr activation time constant, and thus
relatively rapid activation. However, when the early plateau is reduced by ICaL
blockade down to ca. 0 mV, we can see that the time constants become very high
(more than a second), meaning that the IKr does not have enough time
to activate to the steady-state value. At the same time, the same
problem does not manifest in the original ORd model – given its very high
plateau potential, a reduction in plateau caused by ICaL blockade does not
increase IKr activation time constants drastically.
Having understood the problem, what do we do about it? [5].
Going over various publications on IKr activation, I noticed that data underlying ORd IKr have generally fairly slow activation dynamics at the membrane
potentials of -10 to 20 mV. Trying to reduce the activation time constants
ameliorated the problem somewhat, but was not enough. Simply shifting the activation
time constant curves to the left would be another approach we considered, but
to see a sufficient difference, the shift would have to be 15-20 mV, which is a lot.
It would be quite debatable [6] if the model might be called data-driven at that stage without further
justification. I then started exploring more complex Markov models of IKr,
which looked vaguely promising. A complication to the whole process of IKr
explorations was that we had no direct evidence that the ORd IKr
model is problematic - it could be that the rest of the model needed fixing.
The
breakthrough came when my DPhil supervisor Prof. Gil Bub was visiting Oxford,
and with him his PhD supervisor, Prof. Alvin Shrier. Going for a pint of beer
together in the evening, I explained the problem and Prof. Shrier suggested I
should look at the work of Prof. Jamie Vandenberg on IKr and
explained how the single-pulse voltage clamp protocols often fail to
characterize IKr dynamics well, e.g., mixing activation and recovery
from inactivation together.
A question
that might initially look like a sidestep – what might one hope for in the
wildest optimistic dreams? I suggest the following:
- The Vandenberg group published a model of IKr that does not manifest the major reduction in current with reduced plateau following an ICaL blockade.
- The model is easy to incorporate and works without major changes either in itself, or in the rest of the model.
- The paper would also conveniently publish data demonstrating that lowering the AP plateau should not produce a drastic reduction in IKr.
Guess what
– the paper on the Lu-Vandenberg model of IKr does just that! Moments like the
one when I plugged the Lu-Vandenberg model in and it worked out of the box are
the reason why science is so addictive. Looking back at the meeting in the pub,
I guess that corroborates the ages-old wisdom In vino veritas.
Take-home messages
Key points
in my view are:
- The fact that a model is data-driven does not mean that it is bulletproof or that it should be taken as set in stone. For example, as seen in literature on late sodium current and IKr, conventional single-pulse voltage clamp protocol may not produce sufficient information to characterize the behavior of a current when the gating is complex.
- Linking a model’s behavior to a change in a particular current density is a good start of investigation, but seldom the answer. What is described in this blog post started with a fork between good/bad behavior under block of sodium and calcium currents respectively… and the answer was in the replacement of a potassium current formulation. I personally would never discount the possibility that that a computer model gives us answers we like [7] via a problematic mechanism, and so it is important to get to the bottom of the mechanism whenever possible, rather than just associating behaviours with currents.
- Understanding mechanisms is also invaluable in model development whenever an obstacle is reached – it doesn’t give an easy how-to on how to improve the model, but without it, it’s even harder.
[1] The King in Yellow is
present only to make the abbreviation work nicely – nevertheless, it is an
interesting and not that well known old book. Especially if you’re fans of E.
A. Poe or H. P. Lovecraft, give it a try – it’s freely and legally available via Project Gutenberg.
[2] It’s good
to keep in mind the total current amplitude/importance. If there is a relative
change of 20% in a major current like ICaL, it may be a more
important difference than a 50% change in the background calcium current.
[3] That said,
the AP clamp is also not fully bulletproof approach, as the current blocker
used may not be fully specific (it may also block yet unknown
currents/processes), and the current blockade may elicit changes in ionic
concentrations that also modify the cell behavior. I’d say one rather clear
case is when L-type calcium current is estimated as nisoldipine-sensitive
current - it is reasonable to expect that blockade of calcium influx will have
a major effect on the current carried by sodium-calcium exchanger, and this
change will be also reflected in what is measured as the nisoldipine-sensitive
current.
[4] We worked with an intermediate development version of our final
model in this section so far, but for the illustration of the point here, the final ToR-ORd AP shape can be
used, given its similar plateau potentials to the intermediate model.
[5] This may
sound like a common theme of the model development at this stage. However, this
is still a relatively optimistic phase. It’s reasonable to expect spending more
time in the state where one does not understand what the problem is in the
first place, let alone having an idea of the solution.
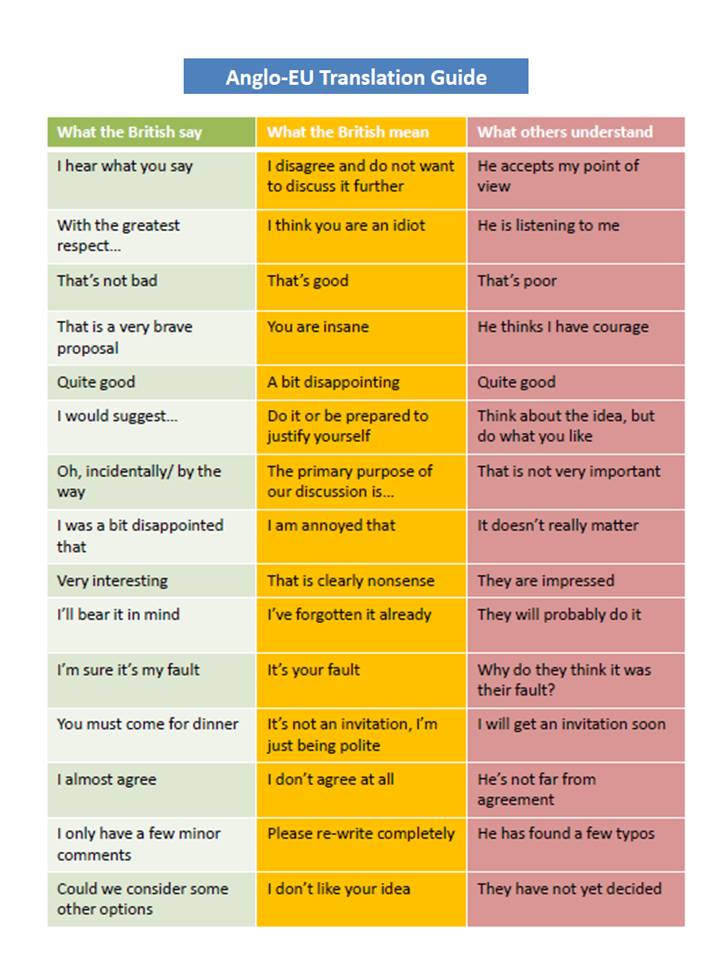
[6] When reading this sentence, please take
into account that I live in the UK for more than 5 years. For more direct styles of communication, the translation to
international English would be more like “No, this really is not data-driven
modelling.” It took me a while to understand that this funny table is an accurate depiction of reality and when I stopped taking it as a joke, I found communication
with native English persons much easier to navigate.
{kind=link}
[7] I suppose
this is linked to a well-known but hard-to-guard-against bias, where one very
critically approaches a problem when things don’t work (looking for new ways to
analyze data, model, etc.), but when the data analysis fits the expectations,
the rigor of criticism suddenly drops and a paper starts being written. But
precisely because it’s a very natural thing and it’s convenient to succumb to this
bias, it’s so important to be aware of it and combat it vigorously. As also seen
in online debates on anything more serious than kittens, I think this is not
about education/knowledge – it’s about attitude and self-criticism.
Comments
Post a Comment